Задание ollbio08101120172018в2
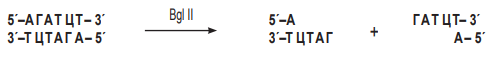 ДНК-лигазой, произойдёт ковалентное соединение фрагментов, соединённых «липкими концами». Это лежит в основе метода получения рекомбинантных ДНК.
При таком сшивании соединение концов одного фрагмента при его длине более 500 нуклеотидных пар происходит в 10 раз чаще, чем соединение концов двух разных фрагментов.
У многих бактерий кроме основной хромосомы присутствуют небольшие дополнительные ДНК, называемые плазмидами. Они представляют собой кольцевые молекулы ДНК, способные к репликации в клетке, и несут гены, отсутствующие в основной хромосоме, например, гены устойчивости к антибиотикам.
Плазмида pСО36 несёт гены устойчивости к эритромицину и ампицилину и состоит из 4200 пар нуклеотидов. Рестриктаза BglII расщепляет эту плазмиду только по гену устойчивости к эритромицину в начале этого гена. В районе расщепления ДНК имеет последовательность нуклеотидов:
ДНК-лигазой, произойдёт ковалентное соединение фрагментов, соединённых «липкими концами». Это лежит в основе метода получения рекомбинантных ДНК.
При таком сшивании соединение концов одного фрагмента при его длине более 500 нуклеотидных пар происходит в 10 раз чаще, чем соединение концов двух разных фрагментов.
У многих бактерий кроме основной хромосомы присутствуют небольшие дополнительные ДНК, называемые плазмидами. Они представляют собой кольцевые молекулы ДНК, способные к репликации в клетке, и несут гены, отсутствующие в основной хромосоме, например, гены устойчивости к антибиотикам.
Плазмида pСО36 несёт гены устойчивости к эритромицину и ампицилину и состоит из 4200 пар нуклеотидов. Рестриктаза BglII расщепляет эту плазмиду только по гену устойчивости к эритромицину в начале этого гена. В районе расщепления ДНК имеет последовательность нуклеотидов:
 Плазмиду обработали рестриктазой BglII до полного расщепления. После этого рестриктазу удалили и смесь фрагментов ДНК обработали ДНК-лигазой. Полученные ДНК смешали с клетками бактерий, не несущих плазмид и неустойчивых к антибиотикам. В результате произошла генетическая трансформация: в часть клеток проникла ДНК плазмиды и изменила их свойства. Полученные клетки высеяли на твёрдую питательную среду, не содержащую антибиотиков. В результате деления каждая клетка образовала
колонию генетически идентичных клеток. Было получено 51366 таких колоний. Клетки из каждой колонии пересеяли на среду, содержащую ампициллин, на которой рост дали 573 колонии. Клетки из колоний,
выросших на ампициллине, пересеяли на среду с эритромицином. На этой среде выросла 51 колония.
Из них выдели плазмидную ДНК, и оказалось что она представлена двумя разными по длине формами, причём каждой колонии был только один вид плазмиды.
А. Какова (в %) эффективность трансформации клеток плазмидной ДНК?
Б. Почему не все колонии, выросшие на ампициллине, дали рост на эритромицине?
В. Как можно объяснить разную длину плазмид в устойчивых к эритромицину колониях?
Г. Сколько всего размерных классов плазмид можно найти в колониях, устойчивых к ампицилину?
Плазмиду обработали рестриктазой BglII до полного расщепления. После этого рестриктазу удалили и смесь фрагментов ДНК обработали ДНК-лигазой. Полученные ДНК смешали с клетками бактерий, не несущих плазмид и неустойчивых к антибиотикам. В результате произошла генетическая трансформация: в часть клеток проникла ДНК плазмиды и изменила их свойства. Полученные клетки высеяли на твёрдую питательную среду, не содержащую антибиотиков. В результате деления каждая клетка образовала
колонию генетически идентичных клеток. Было получено 51366 таких колоний. Клетки из каждой колонии пересеяли на среду, содержащую ампициллин, на которой рост дали 573 колонии. Клетки из колоний,
выросших на ампициллине, пересеяли на среду с эритромицином. На этой среде выросла 51 колония.
Из них выдели плазмидную ДНК, и оказалось что она представлена двумя разными по длине формами, причём каждой колонии был только один вид плазмиды.
А. Какова (в %) эффективность трансформации клеток плазмидной ДНК?
Б. Почему не все колонии, выросшие на ампициллине, дали рост на эритромицине?
В. Как можно объяснить разную длину плазмид в устойчивых к эритромицину колониях?
Г. Сколько всего размерных классов плазмид можно найти в колониях, устойчивых к ампицилину? 📜Теория для решения: Генетический код. Биосинтез белка
Посмотреть решение
Сначала найдём место расщепления плазмиды рестриктазой BglII:
![]()
Таких участков оказывается два. В результате расщепления из плазмиды выщепляется короткий фрагмент:

Остаётся укороченная линейная ДНК, содержащая интактный ген устойчивости к ампицилину и расщеплённый ген устойчивости к эритромицину.
![]()
При сшивании липких концов ДНК-лигазой наиболее часто будут соединяться концы этой молекулы и образовываться кольцо длиной 4163 нуклеотида. Такая ДНК будет сообщать клеткам устойчивость к
ампицилину и не даст устойчивости к эритромицину. Второй фрагмент из-за небольшой длины не может замкнуться в кольцо. Второй вариант лигирования приводит к сшиванию липких концов двух фрагментов. Он происходит примерно в 10 раз реже, а после сшивки вторая пара липких концов скорее всего также, как и исходный фрагмент замкнётся в кольцо. Таких колец из пары фрагментов может образоваться 4 вида: димеры большого фрагмента в двух разных ориентациях (правый конец с левым концом второго фрагмента и левый конец с правым концом второго фрагмента или правый с правым и левый с левым) и соединения большого и малого фрагмента в двух разных ориентациях (вариант исходной плазмиды и инверсия малого фрагмента). Из них только в варианте исходной плазмиды восстанавливается устойчивость к эритромицину.
Линейная молекула, образованная сшиванием двух фрагментов, может присоединить ещё один
фрагмент с ещё в 10 раз меньшей частотой. Такие фрагменты в дальнейшем будут циклизоваться
в плазмиды трёх размеров: из трёх больших фрагментов, из двух больших и одного малого и одного
большого и двух малых. Три малых фрагмента дадут короткую последовательность, которая не сможет
замкнуться в кольцо и существовать в клетке. В каждом размерном классе будет несколько вариантов с
разной ориентацией фрагментов. Только в одном из них восстановится ген устойчивости к эритромицину: правый конец большого фрагмента соединяется с левым концом малого фрагмента, а правый конец малого фрагмента – с левым концом второго большого фрагмента, а оставшиеся концы двух больших фрагментов соединяются с образованием кольцевой плазмиды длиной 8363 пары нуклеотидов. Доля таких молекул будет менее 1% всех плазмид. Вероятность образования плазмид из 4 и более фрагментов ещё на порядок ниже и их обнаружение при данном числе полученных трансформированных клеток нереально.
А. Так как расщепление рестриктазой не затрагивает ген устойчивости к ампицилину, все клетки, в
результате трансформации получившие любую плазмиду, будут устойчивы к ампицилину и вырастут
на среде с этим антибиотиком. Таким образом из 33506 выросших колоний плазмиду получили 578,
выросших на ампицилине. Эффективность трансформации представляет долю трансформированных
клеток от общего их числа, т.е. 573 : 51366 × 100% = 1.12%
Б. На эритромицине могут вырасти только те клетки, в которые попали плазмиды, в которых в
результате лигирования восстановится последовательность нуклеотидов в гене устойчивости к этому
антибиотику, расщеплённому рестриктазой. Остальные плазмиды, полученные по приведённой методике, будут содержать либо ген с выщепленным коротким фрагментом, что приведёт либо к утрате стартового кодона (если обозначенный зелёным цветом кодон является стартовым), либо к сдвигу рамки считывания (т.к. число удалённых нуклеотидов не кратно трём), либо, при инверсии короткого фрагмента, к появлению стоп-кодонов т.е. прекращению синтеза белка. Таким образом большинство полученных плазмид не обеспечат устойчивости к эритромицину.
В. Рост на эритромицине могут обеспечить только плазмиды, несущие восстановленную последовательность гена устойчивости. Такие плазмиды могли образоваться из одного большого и одного малого фрагмента (4200 пар, исходная плазмида)) или из двух больших и одного малого (8363 пары, начало и конец гена из разных копий большого фрагмента).
Г. Получается 1 размер из одного большого фрагмента, два размерных класса из двух фрагментов
и три размерных класса из трёх фрагментов, то есть 6 размерных классов. (В реальности различить
по длине плазмиды, отличающиеся на длину малого фрагмента, т.е. менее чем на 0,5%, невозможно.
Поэтому в эксперименте, например на электрофореграмме, будут видны лишь три размерных класса, соответствующие 1, 2 или 3 копиям большого фрагмента.)